CHAPTER 3 Respiratory Physiology in Infants and Children
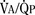 ) for sufficient pulmonary gas exchange. The neonatal adaptation of lung mechanics and respiratory control takes several weeks to complete. Beyond this immediate neonatal period, the infant’s lungs continue to mature at a rapid pace, and postnatal development of the lungs and the thorax surrounding the lungs continues well beyond the first year of life. Respiratory function in infants and toddlers, especially during the first several months of life, as with cardiovascular system and hepatic function, is both qualitatively and quantitatively different from that in older children and adults, and so is their responses to pharmacologic agents, especially anesthetics.
) for sufficient pulmonary gas exchange. The neonatal adaptation of lung mechanics and respiratory control takes several weeks to complete. Beyond this immediate neonatal period, the infant’s lungs continue to mature at a rapid pace, and postnatal development of the lungs and the thorax surrounding the lungs continues well beyond the first year of life. Respiratory function in infants and toddlers, especially during the first several months of life, as with cardiovascular system and hepatic function, is both qualitatively and quantitatively different from that in older children and adults, and so is their responses to pharmacologic agents, especially anesthetics.Development of the respiratory system
Prenatal Development of the Lungs
The morphologic development of the human lung is seen as early as several weeks into the embryonic period and continues well into the first decade of postnatal life and beyond (Fig. 3-1). The fetal lungs begin to form within the first several weeks of the embryonic period, when the fetus is merely 3 mm in length. A groove appears in the ventral aspect of the foregut, creating a small pouch. The outgrowth of the endodermal cavity, with a mass of surrounding mesenchymal tissue, projects into the pleuroperitoneal cavity and forms lung buds. The future alveolar membranes and mucous glands are derived from the endoderm, whereas the cartilage, muscle, elastic tissue, and lymph vessels originate from the mesenchymal elements surrounding the lung buds (Emery, 1969).
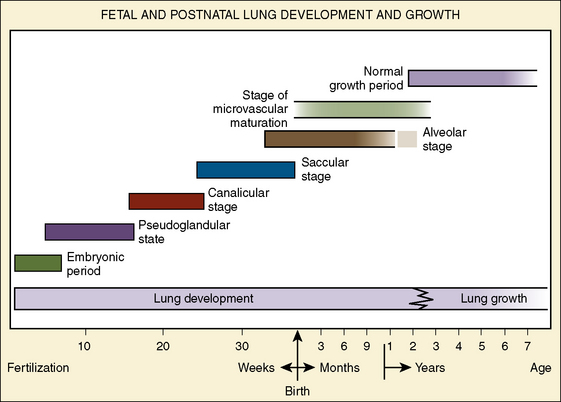
During the pseudoglandular period, which extends until 17 weeks’ gestation, the budding of the bronchi and lung growth rapidly take place, forming a loose mass of connective tissue. The morphologic development of the human lung is illustrated in Figure 3-2. By 16 weeks’ gestation, preacinar branching of the airways (down to the terminal bronchioli) is complete (Reid, 1967). A disturbance of the free expansion of the developing lung during this stage, as occurs with diaphragmatic hernia, results in hypoplasia of the airways and lung tissue (Areechon and Reid, 1963). During the canalicular period, in midgestation, the future respiratory bronchioli develop as the relative amount of connective tissue diminishes. Capillaries grow adjacent to the respiratory bronchioli, and the whole lung becomes more vascular (Emery, 1969).
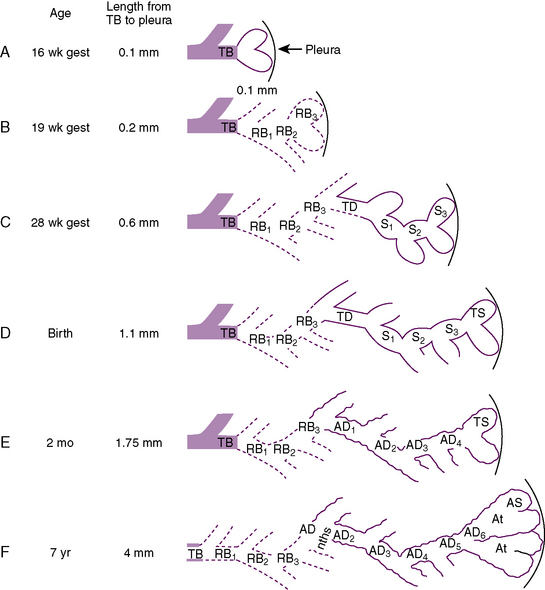
At about 24 weeks’ gestation, the lung enters the terminal sac period, which is characterized by the appearance of clusters of terminal air sacs, termed saccules, with flattened epithelium (Hislop and Reid, 1974). These saccules are large and irregular with thick septa and have few capillaries in comparison with the adult alveoli (Boyden, 1969). At about 26 to 28 weeks’ gestation, proliferation of the capillary network surrounding the terminal air spaces becomes sufficient for pulmonary gas exchange (Potter, 1961). These morphologic developments may occur earlier in some premature infants (born at 24 to 25 weeks’ gestation) who have survived through neonatal intensive care. Starting at 28 weeks’ gestation, air space wall thickness decreases rapidly. From this period onward toward term, there is further lengthening of saccules with possible growth of additional generations of air spaces. Some mammalian species, such as the rat, have no mature alveoli at birth (Burri, 1974). In contrast, alveolar development from saccules begins in some human fetuses as early as 32 weeks’ gestation, but alveoli are not uniformly present until 36 weeks’ gestation (Langston et al., 1984). Most alveolar formation in humans takes place postnatally during the first 12 to 18 months of postnatal life, and development of respiratory bronchioles by transformation of preexisting terminal airways does not take place until after birth (Langston et al., 1984).
The fetal lung produces a large quantity of liquid, which expands the airways while the larynx is closed. This expansion produces the growth factor, such as human bombesin, and helps to stimulate lung growth and development (Sunday et al., 1988). The lung fluid is periodically expelled into the uterine cavity and contributes about one third of the total amniotic fluid. Prenatal ligation or occlusion of the trachea was tried in the 1990s with some success for the treatment of the fetus with congenital diaphragmatic hernia (Harrison et al., 1993). This treatment causes the expansion of the fetal airways and results in an accelerated growth of the otherwise hypoplastic lung.
The type II pneumocytes, which produce pulmonary surfactant that forms the alveolar lining layer, reduces surface tension and stabilizes air spaces after air breathing, appear at about 24 to 26 weeks’ gestation but occasionally as early as 20 weeks (Spear et al., 1969; Lauweryns, 1970). Idiopathic (or infantile) respiratory distress syndrome (IRDS), also known as hyaline membrane disease (HMD), which occurs in premature infants, is caused by the immaturity of the lungs with their insufficient pulmonary surfactant production and their inactivation by plasma proteins exudating onto the alveolar surface (see Surface Activity and Pulmonary Surfactant).
Experimental evidence from animals indicates that certain pharmacologic agents such as cortisol and thyroxin administered to the mother or directly to the fetus accelerate the maturation of the lungs, resulting in the early appearance of type II pneumocytes and surfactant (deLemos et al., 1970; Motoyama et al., 1971; Wu et al., 1973; Smith and Bogues, 1982; Rooney, 1985). Liggins and Howie (1972) reported accelerated maturation of human fetal lungs after the administration of corticosteroids to mothers 24 to 48 hours before the delivery of premature babies. Despite initial concern that steroids might potentially be toxic to other organs of the fetus, particularly to the development of the central nervous system, prenatal glucocorticoid therapy has been used widely since the 1980s to induce lung maturation and surfactant synthesis in mothers at risk of premature delivery (Avery, 1984; Avery et al., 1986).
Neonatal respiratory adaptation
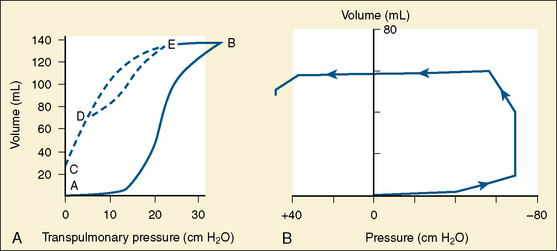
To introduce air into the fluid-filled lungs at birth, the newborn infant must overcome large surface force with the first few breaths. Usually a negative pressure of 30 cm H2O is necessary to introduce air into the fluid-filled lungs. In some normal full-term infants, even with sufficient surfactant, a force of as much as −70 cm H2O or more must be exerted to overcome the surface force (Fig. 3-3) (Karlberg et al., 1962). Usually fluid is rapidly expelled via the upper airways. The residual fluid leaves the lungs through the pulmonary capillaries and lymphatic channels over the first few days of life, and changes in compliance parallel this time course. All changes are delayed in the premature infant.
As the lungs expand with air, pulmonary vascular resistance decreases dramatically and pulmonary blood flow increases markedly, thus allowing gas exchange between alveolar air and pulmonary capillaries to occur. Changes in Po2, Pco2 (P stands for partial pressure), and pH are largely responsible for the dramatic decrease in pulmonary vascular resistance (Cook et al., 1963). The resultant large increases in pulmonary blood flow and the increase in left atrial pressure with a decrease in right atrial pressure reverse the pressure gradient across the atria and close (initially functionally and eventually anatomically) the foramen ovale, a right-to-left one-way valve. With these adjustments, the cardiopulmonary system approaches adult levels of ventilation/perfusion (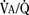 balance within a few days (Nelson et al., 1962, 1963). The process of expansion of the lungs during the first few hours of life and the resultant circulatory adaptation for establishing pulmonary gas exchange are greatly influenced by the adequacy of pulmonary surfactant. It should be remembered that these changes are delayed in immature newborns.
balance within a few days (Nelson et al., 1962, 1963). The process of expansion of the lungs during the first few hours of life and the resultant circulatory adaptation for establishing pulmonary gas exchange are greatly influenced by the adequacy of pulmonary surfactant. It should be remembered that these changes are delayed in immature newborns.
Postnatal Development of the Lungs and Thorax
The development and growth of the lungs and surrounding thorax continue with amazing speed during the first year of life. Although the formation of the airway system all the way to the terminal bronchioles is complete by 16 weeks’ gestation, alveolar formation begins only at about 36 weeks’ gestation. At birth, the number of terminal air sacs (most of which are saccules) is between 20 and 50 million, only one tenth that of fully grown lungs of the child. Most postnatal development of alveoli from primitive saccules occurs during the first year and is essentially completed by 18 months of age (Langston et al., 1984). The morphologic and physiologic development of the lungs, however, continues throughout the first decade of life (Mansell et al., 1972).
In the neonate, static (elastic) recoil pressure of the lungs is very low (i.e., compliance, normalized for volume, is unusually high), which is not dissimilar to that of geriatric or emphysematous lungs, because the elastic fibers do not develop until the postnatal period (whereas elastic fibers in geriatric lungs are brittle and not functional) (Mansel et al., 1972; Fagan, 1976; Bryan and Wohl, 1986). In addition, the elastic recoil pressure of the infant’s thorax (chest wall) is extremely low because of its compliant cartilaginous rib cage with poorly developed thoracic muscle mass, which does not add rigidity. These unique characteristics make infants more prone to lung collapse, especially under general anesthesia when inspiratory muscles are markedly relaxed (see maintenance of FRC below). Throughout infancy and childhood, static recoil pressure of the lungs and thorax steadily increases (compliance, normalized for volume, decreases) toward normal values for young adults (Zapletal et al., 1971; Motoyama, 1977).
The actual size of the airway from the larynx to the bronchioles in infants and children, of course, is much smaller than in adolescents and adults, and flow resistance in absolute terms is extremely high. When normalized for lung volume or body size, however, infants’ airway size is relatively much larger; airway resistance is much lower than in adults (Polgar, 1967; Motoyama, 1977; Stocks and Godfrey, 1977). Infants and toddlers, however, are more prone to severe obstruction of the upper and lower airways because their absolute (not relative) airway diameters are much smaller than those in adults. As a consequence, relatively mild airway inflammation, edema, or secretions can lead to far greater degrees of airway obstruction than in adults (e.g., as with subglottic croup [laryngotracheobronchitis] or acute supraglottitis [epiglottitis]).
Prenatal Development of Breathing
Respiratory rhythmogenesis occurs long before parturition. Dawes and others (1970) were the first to demonstrate “breathing” activities with rhythmic diaphragmatic contractions in the fetal lamb. They found it to be episodic and highly variable in frequency. Boddy and Robinson (1971) recorded movement of the human fetal thorax with an ultrasound device and interpreted this as evidence of fetal breathing. Later studies have shown that during the last 10 weeks of pregnancy, fetal breathing is present approximately 30% of the time (Patrick et al., 1980). The breathing rate in the fetus at 30 to 31 weeks’ gestation is higher (58 breaths/min) than that in the near-term fetus (47 breaths/min). A significant increase in fetal breathing movements occurs 2 to 3 hours after a maternal meal and is correlated with the increase in the maternal blood sugar level (Patrick et al., 1980).
Spontaneous breathing movements in the fetus occur only during active, or rapid eye movement (REM), sleep and with low-voltage electrocortical activity, and they appear to be independent of the usual chemical and nonchemical stimuli of postnatal breathing (Dawes et al., 1972; Jansen and Chernick, 1983). Later studies, however, have clearly shown that the fetus can respond to chemical stimuli known to modify breathing patterns postnatally (Dawes et al., 1982; Jansen et al., 1982; Rigatto et al., 1988, 1992). In contrast, hypoxemia in the fetus abolishes, rather than stimulates, breathing movements. This may be related to the fact that hypoxemia diminishes the incidence of REM sleep (Boddy et al., 1974). It appears that normally low arterial oxygen tension, or Pao2 (19 to 23 mm Hg), in the fetus is a normal mechanism inhibiting breathing activities in utero (Rigatto, 1992). Severe hypoxia induces gasping, which is independent of the peripheral chemoreceptors and apparently independent of rhythmic fetal breathing (Jansen and Chernick, 1974).
The near-term fetus is relatively insensitive to Paco2 changes. Extreme hypercapnia (Paco2 greater than 60 mm Hg) in the fetal lamb, however, can induce rhythmic breathing movement that is preceded by a sudden activation of inspiratory muscle tone with expansion of the thorax and inward movement (inspiration) of amniotic fluid, as much as 30 to 40 mL/kg (an apparent increase in functional residual capacity [FRC]) (Motoyama, unpublished observation). When Pao2 was reduced, breathing activities ceased, and there was a reversal of the sequence of events noted above (i.e., relaxation of the thorax, decreased FRC as evidenced by outward flow of amniotic fluid) (Motoyama, 2001).
The Hering-Breuer (inflation) reflex is present in the fetus. Distention of the lungs by saline infusion slows the frequency of breathing (Dawes et al., 1982). Transection of the vagi, however, does not change the breathing pattern (Dawes, 1974).
Maternal ingestion of alcoholic beverages abolishes human fetal breathing for up to 1 hour. Fetal breathing movement is also abolished by maternal cigarette smoking. These effects may be related to fetal hypoxemia resulting from changes in placental circulation Jansen and Chernick, 1983). It is not clear why the fetus must “breathe” in utero, when gas exchange is handled by the placental circulation. Dawes (1974) suggested that fetal breathing might represent “prenatal practice” to ensure that the respiratory system is well developed and ready at the moment of birth. Another reason may be that the stretching of the airways and lung parenchyma is an important stimulus for lung development; bilateral phrenic nerve sectioning in the fetal lamb results in hypoplasia of the lungs (Alcorn et al., 1980).
Perinatal Adaptation of Breathing
During normal labor and vaginal delivery, the human fetus goes through a period of transient hypoxia, hypercapnia, and acidemia. The traditional view of the mechanism of the onset of breathing at birth until the 1980s was that the transient fetal asphyxia stimulates the chemoreceptors and produces gasping, which is followed by rhythmic breathing at birth that is aided by thermal, tactile, and other sensory stimuli. Subsequent studies have challenged this concept (Chernick et al., 1975; Baier et al., 1990; Rigatto, 1992). The current concept regarding the mechanism of continuous neonatal breathing is summarized in Box 3-1.
Box 3-1 Mechanism of Continuous Neonatal Breathing
Once the newborn has begun rhythmic breathing, ventilation is adjusted to achieve a lower Paco2 than is found in older children and adults (Table 3-1). The reason for this difference is not clear but most likely is related to a poor buffering capacity in the neonate and a ventilatory compensation for metabolic acidosis. The Pao2 of the infant approximates the adult level within a few weeks of birth (Nelson, 1976).
Control of breathing
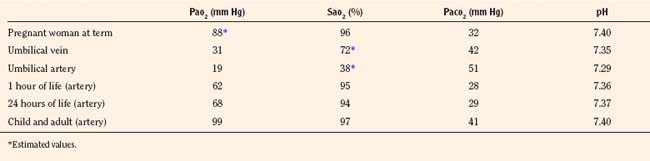
Rhythmic contraction of the respiratory muscles is governed by the respiratory centers in the brainstem and tightly regulated by feedback systems so as to match the level of ventilation to metabolic needs (Fig. 3-4) (Cherniack and Pack, 1988). These feedback mechanisms include central and peripheral chemoreceptors, stretch receptors in the airways and lung parenchyma via the vagal afferent nerves, and segmental reflexes in the spinal cord provided by muscle spindles (Cherniack and Pack, 1988). The control of breathing comprises neural and chemical controls that are closely interrelated.
Neural Control of Breathing
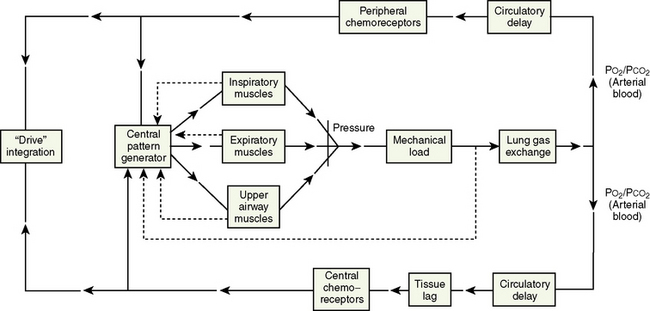
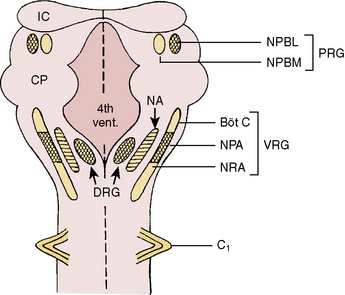
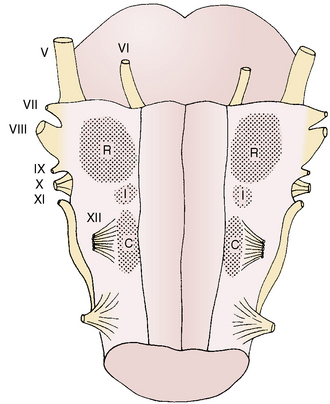
Respiratory neurons in the medulla have inherent rhythmicity even when they are separated from the higher levels of the brainstem. In the cat, respiratory neurons are concentrated in two bilaterally symmetric areas in the medulla near the level of the obex. The dorsal respiratory group of neurons (DRG) is located in the dorsomedial medulla just ventrolateral to the nucleus tractus solitarius and contains predominantly inspiratory neurons. The ventral respiratory group of neurons (VRG), located in the ventrolateral medulla, consists of both inspiratory and expiratory neurons (Fig. 3-5) (von Euler, 1986; Tabatabai and Behnia, 1995; Berger, 2000).
Dorsal Respiratory Group of Neurons
The DRG is spatially associated with the tractus solitarius, which is the principal tract for the ninth and tenth cranial (glossopharyngeal and vagus) nerves. These nerves carry afferent fibers from the airways and lungs, heart, and peripheral arterial chemoreceptors. The DRG may constitute the initial intracranial site for processing some of these visceral sensory afferent inputs into a respiratory motor response (Berger, 2000).
On the basis of lung inflation, three types of neurons have been recognized in the DRG: type Iα (I stands for inspiratory), type Iβ, and pump (P) cells. Type Iα is inhibited by lung inflation (Cohen, 1981a). The axons of these neurons project to both the phrenic and the external (inspiratory) intercostal motoneurons of the spinal cord. Some type Iα neurons have medullary collaterals that terminate among the inspiratory and expiratory neurons of the ipsilateral VRG (Merrill, 1970).
The second type, Iβ, is excited by lung inflation and receives synaptic inputs from pulmonary stretch receptors. There is controversy as to whether Iβ axons project into the spinal cord respiratory neurons; the possible functional significance of such spinal projections is unknown. Both Iα and Iβ neurons receive excitatory inputs from the central pattern generator (or central inspiratory activity) for breathing, so that when lung inflation is terminated or the vagi in the neck are cut, the rhythmic firing activity of these neurons continues (Cohen, 198la, 1981b; Feldman and Speck, 1983).
The third type of neurons in the DRG receives no input from the central pattern generator. The impulse of these neurons, the P cells, closely follows lung inflation during either spontaneous or controlled ventilation (Berger, 1977). The P cells are assumed to be relay neurons for visceral afferent inputs (Berger, 2000).
The excitation of Iβ neurons by lung inflation is associated with the shortening of inspiratory duration. The Iβ neurons appear to promote inspiration-to-expiration phase-switching by inhibiting Iα neurons. This network seems to be responsible for the Hering-Breuer reflex inhibition of inspiration by lung inflation (Cohen, 198la, 1981b; von Euler, 1986, 1991).
The DRG thus functions as an important primary and possibly secondary relay site for visceral sensory inputs via glossopharyngeal and vagal afferent fibers. Because many of the inspiratory neurons in the DRG project to the contralateral spinal cord and make excitatory connections with phrenic motoneurons, the DRG serves as a source of inspiratory drive to phrenic and possibly to external intercostal motoneurons (Berger, 2000).
Ventral Respiratory Group of Neurons
The VRG extends from the rostral to the caudal end of the medulla and has three subdivisions (Fig. 3-5). The Bötzinger complex, located in the most rostral part of the medulla in the vicinity of the retrofacial nucleus, contains mostly expiratory neurons (Lipski and Merrill, 1980; Merrill et al., 1983). These neurons send inhibitory signals to DRG and VRG neurons and project into the phrenic motoneurons of the spinal cord, causing its inhibition (Bianchi and Barillot, 1982; Merrill et al., 1983). The physiologic significance of these connections may be to ensure inspiratory neuronal silence during expiration (reciprocal inhibition) and to contribute to the “inspiratory off-switch” mechanism.
The nucleus ambiguus (NA) and nucleus paraambigualis (NPA), lying side by side, occupy the middle portion of the VRG. Axons of the respiratory motoneurons originating from the NA project along with other vagal efferent fibers and innervate the laryngeal abductor (inspiratory) and adductor (expiratory) muscles via the recurrent laryngeal nerve (Barillot and Bianchi, 1971; Bastel and Lines, 1975). The NPA contains mainly inspiratory (Iγ) neurons, which respond to lung inflation in a manner similar to that of Iα neurons. The axons of these neurons project both to phrenic and external (inspiratory) intercostal motoneuron pools in the spinal cord. The nucleus retroambigualis (NRA) occupies the caudal part of the VRG and contains expiratory neurons whose axons project into the spinal motoneuron pools for the internal (expiratory) intercostal and abdominal muscles (Merrill, 1970; Miller et al., 1985).
The inspiratory neurons of the DRG send collateral fibers to the inspiratory neurons of the NPA in the VRG. These connections may provide the means for ipsilateral synchronization of the inspiratory activity between the neurons in the DRG and those in the VRG (Merrill, 1979, 1983). Furthermore, axon collaterals of the inspiratory neurons of the NPA on one side project to the inspiratory neurons of the contralateral NPA, and vice versa. These connections may be responsible for the bilateral synchronization of the medullary inspiratory motoneuron output, as evidenced by synchronous bilateral phrenic nerve activity (Merrill, 1979, 1983).
Pontine Respiratory Group of Neurons
In the dorsolateral portion of the rostral pons, both inspiratory and expiratory neurons have been found. Inspiratory neuronal activity is concentrated ventrolaterally in the region of the nucleus parabrachialis lateralis (NPBL). The expiratory activity is centered more medially in the vicinity of the nucleus parabrachialis medialis (NPBM) (Fig. 3-5) (Cohen, 1979; Mitchell and Berger, 1981). The respiratory neurons of these nuclei are referred to as the pontine respiratory group (PRG), which was, and sometimes still is, called the pneumotaxic center, although the term is generally considered obsolete (Feldman, 1986). There are reciprocal projections between the PRG neurons and the DRG and VRG neurons in the medulla. Electrical stimulation of the PRG produces rapid breathing with premature switching of respiratory phases, whereas transaction of the brainstem at a level caudal to the PRG prolongs inspiratory time (Cohen, 1971; Feldman and Gautier, 1976). Bilateral cervical vagotomies produce a similar pattern of slow breathing with prolonged inspiratory time; a combination of PRG lesions and bilateral vagotomy in the cat results in apneusis (apnea with sustained inspiration) or apneustic breathing (slow rhythmic respiration with marked increase end inspiratory hold) (Feldman and Gaultier, 1976; Feldman, 1986). The PRG probably plays a secondary role in modifying the inspiratory off-switch mechanism (Gautier and Bertrand, 1975; von Euler and Trippenbach, 1975).
Respiratory Rhythm Generation
Rhythmic breathing in mammals can occur in the absence of feedback from peripheral receptors. Because transection of the brain rostral to the pons or high spinal transection has little effect on the respiratory pattern, respiratory rhythmogenesis apparently takes place in the brainstem. The PRG, DRG, and VRG have all been considered as possible sites of the central pattern generator, although its exact location is still unknown (Cohen, 1981b; von Euler, 1983, 1986). A study with an in vitro brainstem preparation of neonatal rats has indicated that respiratory rhythm is generated in the small area in the ventrolateral medulla just rostral to the Bötzinger complex (pre-Bötzinger complex), which contains pacemaker neurons (Smith et al., 1991).
The pre-Bötzinger complex contains a group of neurons that is responsible for respiratory rhythmogenesis (Smith et al., 1991; Pierrefiche et al., 1998; Rekling and Feldman, 1998). Although the specific cellular mechanism responsible for rhythmogenesis is not known, two possible mechanisms have been proposed (Funk and Feldman, 1995; Ramirez and Richter, 1996). One hypothesis is that the pacemaker neurons possess intrinsic properties associated with various voltage- and time-dependent ion channels that are responsible for rhythm generation. Rhythmic activity in these neurons may depend on the presence of an input system that may be necessary to maintain the neuron’s membrane potential in a range in which the voltage-dependent properties of the cell’s ion channels result in rhythmic behavior. The network hypothesis is the alternative model in which the interaction between the neurons produces respiratory rhythmicity, such as reciprocal inhibition between inhibitory and excitatory neurons and recurrent excitation within any population of neurons (Berger, 2000). The output of this central pattern generator is influenced by various inputs from chemoreceptors (central and peripheral), mechanoreceptors (e.g., pulmonary receptors and muscle and joint receptors), thermoreceptors (central and peripheral), nociceptors, and higher central structures (such as the PRG). The function of these inputs is to modify the breathing pattern to meet and adjust to ever-changing metabolic and behavioral needs (Smith et al., 1991).
Airway and Pulmonary Receptors
Upper Airway Receptors
Stimulation of receptors in the nose can produce sneezing, apnea, changes in bronchomotor tone, and the diving reflex, which involves both the respiratory and the cardiovascular systems. Stimulation of the epipharynx causes the sniffing reflex, a short, strong inspiration to bring material (mucus, foreign body) in the epipharynx into the pharynx to be swallowed or expelled. The major role of receptors in the pharynx is associated with swallowing. It involves the inhibition of breathing, closure of the larynx, and coordinated contractions of pharyngeal muscles (Widdicombe, 1985; Nishino, 1993; Sant’Ambrogio et al., 1995).
The larynx has a rich innervation of receptors. The activation of these receptors can cause apnea, coughing, and changes in the ventilatory pattern (Widdicombe, 1981, 1985). These reflexes, which influence both the patency of the upper airway and the breathing pattern, are related to transmural pressure and air flow. Based on single-fiber action-potential recordings from the superior laryngeal nerve in the spontaneously breathing dog preparation in which the upper airway is isolated from the lower airways, three types of receptors have been identified: pressure receptors (most common, about 65%), “drive” (or irritant) receptors (stimulated by upper airway muscle activities), and flow or cold receptors (Sant’Ambrogio et al., 1983; Fisher et al., 1985). The laryngeal flow receptors show inspiratory modulation with room air breathing but become silent when inspired air temperature is raised to body temperature and 100% humidity or saturation (Sant’Ambrogio et al., 1985). The activity of pressure receptors increases markedly with upper airway obstruction (Sant’Ambrogio et al., 1983).
Tracheobronchial and Pulmonary Receptors
Three major types of tracheobronchial and pulmonary receptors have been recognized: slowly adapting (pulmonary stretch) receptors and rapidly adapting (irritant or deflation) receptors, both of which lead to myelinated vagal afferent fibers and unmyelinated C-fiber endings (J-receptors). Excellent reviews on pulmonary receptors have been published (Pack, 1981; Widdicombe, 1981; Sant’Ambrogio, 1982; Coleridge and Coleridge, 1984).
Slowly adapting (pulmonary stretch) receptors
Slowly adapting (pulmonary stretch) receptors (SARs) are mechanoreceptors that lie within the submucosal smooth muscles in the membranous posterior wall of the trachea and central airways (Bartlett et al., 1976). A small proportion of the receptors are located in the extrathoracic upper trachea (Berger, 2000). SARs are activated by the distention of the airways during lung inflation and inhibit inspiratory activity (Hering-Breuer inflation reflex), whereas they show little response to steady levels of lung inflation. The Hering-Breuer reflex also produces dilation of the upper airways from the larynx to the bronchi. Although SARs are predominantly mechanoreceptors, hypocapnia stimulates their discharge, and hypercapnia inhibits it (Pack, 1981). In addition, SARs are thought to be responsible for the accelerated heart rate and systemic vasoconstriction observed with moderate lung inflation (Widdicombe, 1974). These effects are abolished by bilateral vagotomy.
Studies by Clark and von Euler (1972) have demonstrated the importance of the inflation reflex in adjusting the pattern of ventilation in the cat and the human. In cats anesthetized with pentobarbital, inspiratory time decreases as tidal volume increases with hypercapnia, indicating the presence of the inflation reflex in the normal tidal volume range. Clark and von Euler demonstrated an inverse hyperbolic relationship between the tidal volume and inspiratory time. In the adult human, inspiratory time is independent of tidal volume until the latter increases to about twice the normal tidal volume, when the inflation reflex appears (Fig. 3-6). In the newborn, particularly the premature newborn, the inflation reflex is present in the eupneic range for a few months (Olinsky et al., 1974).
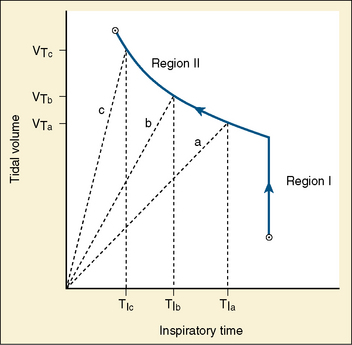
Apnea, commonly observed in adult patients at the end of surgery and anesthesia with the endotracheal tube cuff still inflated, may be related to the inflation reflex, because the trachea has a high concentration of stretch receptors (Bartlett et al., 1976; Sant’Ambrogio, 1982). Deflation of the cuff promptly restores rhythmic spontaneous ventilation.
Rapidly adapting (irritant) receptors
Rapidly adapting (irritant) receptors (RARs) are located superficially within the airway epithelial cells, mostly in the region of the carina and the large bronchi (Pack, 1981; Sant’Ambrogio, 1982). RARs respond to both mechanical and chemical stimuli. In contrast to SARs, RARs adapt rapidly to large lung inflation, distortion, or deflation, thus possessing marked dynamic sensitivity (Pack, 1981). RARs are stimulated by cigarette smoke, ammonia, and other irritant gases including inhaled anesthetics, with significant interindividual variability (Sampson and Vidruk, 1975). RARs are stimulated more consistently by histamine and prostaglandins, suggesting their role in response to pathologic states (Coleridge et al., 1976; Sampson and Vidruk, 1977; Vidruk et al., 1977; Berger, 2000). The activation of RARs in the large airways may be responsible for various reflexes, including coughing, bronchoconstriction, and mucus secretion. Stimulation of RARs in the periphery of the lungs may produce hyperpnea. Because RARs are stimulated by deflation of the lungs to produce hyperpnea in animals, they are considered to play an important role in the Hering-Breuer deflation reflex (Sellick and Widdicombe, 1970). This reflex, if it exists in humans, may partly account for increased respiratory drive when the lung volume is abnormally decreased, as in premature infants with IRDS and in pneumothorax.
When vagal conduction is partially blocked by cold, inflation of the lung produces prolonged contraction of the diaphragm and deep inspiration instead of inspiratory inhibition. This reflex, the paradoxical reflex of Head, is most likely mediated by RARs. It may be related to the complementary cycle of respiration, or the sigh mechanism, that functions to reinflate and reaerate parts of the lungs that have collapsed because of increased surface force during quiet, shallow breathing (Mead and Collier, 1959). In the newborn, inflation of the lungs initiates gasping. This mechanism, which was considered to be analogous to the paradoxical reflex of Head, may help to inflate unaerated portions of the newborn lung (Cross et al., 1960).
C-Fiber endings
Most afferent axons arising from the lungs, heart, and other abdominal viscera are slow conducting (slower than 2.5 m/sec), unmyelinated vagal fibers (C-fibers). Extensive studies by Paintal (1973) have suggested the presence of receptors supposedly located near the pulmonary or capillary wall (juxtapulmonary capillary or J-receptors) innervated by such C-fibers. C-fiber endings are stimulated by pulmonary congestion, pulmonary edema, pulmonary microemboli, and irritant gases such as anesthetics. Such stimulation causes apnea followed by rapid, shallow breathing, hypotension, and bradycardia. Stimulation of J-receptors also produces bronchoconstriction and increases mucus secretion. All these responses are abolished by bilateral vagotomy. In addition, stimulation of C-fiber endings can provoke severe reflex contraction of the laryngeal muscles, which may be partly responsible for the laryngospasm observed during induction of anesthesia with isoflurane or halothane.
In addition to receptors within the lung parenchyma (pulmonary C-fiber endings), there appear to be similar nonmyelinated nerve endings in the bronchial wall (bronchial C-fiber endings) (Coleridge and Coleridge, 1984). Both chemical and, to a lesser degree, mechanical stimuli excite these bronchial C-fiber endings. They are also stimulated by endogenous mediators of inflammation, including histamine, prostaglandins, serotonin, and bradykinin. Such stimulation may be a mechanism of C-fiber involvement in disease states such as pulmonary edema, pulmonary embolism, and asthma (Coleridge and Coleridge, 1984).
The inhalation of irritant gases or particles causes a sensation of tightness or distress in the chest, probably caused by its activation of pulmonary receptors. The pulmonary receptors may contribute to the sensation of dyspnea in lung congestion, atelectasis, and pulmonary edema. Bilateral vagal blockade in patients with lung disease abolished dyspneic sensation and increased breath-holding time (Noble et al., 1970).
Chest-Wall Receptors
The chest-wall muscles, including the diaphragm and the intercostal muscles, contain various types of receptors that can produce respiratory reflexes. This subject has been reviewed extensively (Newsom-Davis, 1974; Duron, 1981). The two types of receptors that have been most extensively studied are muscle spindles, which lie parallel to the extrafusal muscle fibers, and the Golgi tendon organs, which lie in series with the muscle fibers (Berger, 2000).
Intercostal muscles have a density of muscle spindles comparable with those of other skeletal muscles. The arrangement of muscle spindles is appropriate for the respiratory muscle load-compensation mechanism (Berger, 2000). By comparison with the intercostal muscles, the diaphragm has a very low density of muscle spindles and is poorly innervated by the γ-motoneurons. Reflex excitation of the diaphragm, however, can be achieved via proprioceptive excitation within the intercostal system (Decima and von Euler, 1969).
Golgi tendon organs are located at the point of insertion of the muscle fiber into its tendon and, like muscle spindles, are a slowly adapting mechanoreceptor. Activation of the Golgi tendon organs inhibits the homonymous motoneurons, possibly preventing the muscle from being overloaded (Berger, 2000). In the intercostal muscles, fewer Golgi tendon organs are present than muscle spindles, whereas the ratio is reversed in the diaphragm.
Chemical Control of Breathing
Regulation of alveolar ventilation and maintenance of normal arterial Pco2, pH, and Po2 are the principal functions of the medullary and peripheral chemoreceptors (Leusen, 1972).
Central Chemoreceptors
The medullary, or central, chemoreceptors, located near the surface of the ventrolateral medulla, are anatomically separated from the medullary respiratory center (Fig. 3-7). They respond to changes in hydrogen ion concentration in the adjacent cerebrospinal fluid rather than to changes in arterial Pco2 or pH (Pappenheimer et al., 1965). Since CO2 rapidly passes through the blood-brain barrier into the cerebrospinal fluid, which has poor buffering capacity, the medullary chemoreceptors are readily stimulated by respiratory acidemia. In contrast, ventilatory responses of the medullary chemoreceptors to acute metabolic acidemia and alkalemia are limited because changes in the hydrogen ion concentration in arterial blood are not rapidly transmitted to the cerebrospinal fluid. In chronic acid-base disturbances, the pH of cerebrospinal fluid (and presumably that of interstitial fluid) surrounding the medullary chemoreceptors is generally maintained close to the normal value of about 7.3 regardless of arterial pH (Mitchell et al., 1965). Under these circumstances, ventilation becomes more dependent on the hypoxic response of peripheral chemoreceptors.
Peripheral Chemoreceptors
The carotid bodies, located near the bifurcation of the common carotid artery, react rapidly to changes in Pao2 and pH. Their contribution to the respiratory drive amounts to about 15% of resting ventilation (Severinghaus, 1972). The carotid body has three types of neural components: type I (glomus) cells, presumably the primary site of chemotransduction; type II (sheath) cells; and sensory nerve fibers (McDonald, 1981). Sensory nerve fibers originate from terminals in apposition to the glomus cells, travel via the carotid sinus nerve to join the glossopharyngeal nerve, and then enter the brainstem. The sheath cells envelop both the glomus cells and the sensory nerve terminals. A variety of neurochemicals have been found in the carotid body, including acetylcholine, dopamine, substance P, enkephalins, and vasoactive intestinal peptide. The exact functions of these cell types and the mechanisms of chemotransduction and the specific roles of these neurochemicals have not been well established (Berger, 2000).
The carotid bodies are perfused with extremely high levels of blood flow and respond rapidly to an oscillating Pao2 rather than a constant Pao2 at the same mean values (Dutton et al., 1964; Fenner et al., 1968). This mechanism may be partly responsible for hyperventilation during exercise.
The primary role of peripheral chemoreceptors is their response to changes in arterial Po2. Moderate to severe hypoxemia (Pao2 less than 60 mm Hg) results in a significant increase in ventilation in all age groups except for newborn, particularly premature, infants, whose ventilation is decreased by hypoxemia (Dripps and Comroe, 1947; Rigatto et al., 1975b). Peripheral chemoreceptors are also partly responsible for hyperventilation in hypotensive patients. Respiratory stimulation is absent in certain states of tissue hypoxia, such as moderate to severe anemia and carbon monoxide poisoning; despite a decrease in oxygen content, Pao2 in the carotid bodies is maintained near normal levels, so that the chemoreceptors are not stimulated.
In acute hypoxemia, the ventilatory response via the peripheral chemoreceptors is partially opposed by hypocapnia, which depresses the medullary chemoreceptors. When a hypoxemic environment persists for a few days, for example, during an ascent to high altitude, ventilation increases further as cerebrospinal fluid bicarbonate decreases and pH returns toward normal (Severinghaus et al., 1963). However, later studies demonstrated that the return of cerebrospinal fluid pH toward normal is incomplete, and a secondary increase in ventilation precedes the decrease in pH, indicating that some other mechanisms are involved (Bureau and Bouverot, 1975; Foster et al., 1975). In chronic hypoxemia that lasts for a number of years, the carotid bodies initially exhibit some adaptation to hypoxemia and then gradually lose their hypoxic response. In people native to high altitudes, the blunted response of carotid chemoreceptors to hypoxemia takes 10 to 15 years to develop and is sustained thereafter (Sorensen and Severinghaus, 1968; Lahiri et al., 1978). In cyanotic heart diseases, the hypoxic response is lost much sooner but returns after surgical correction of the right-to-left shunts (Edelman et al., 1970).
Response to Carbon Dioxide
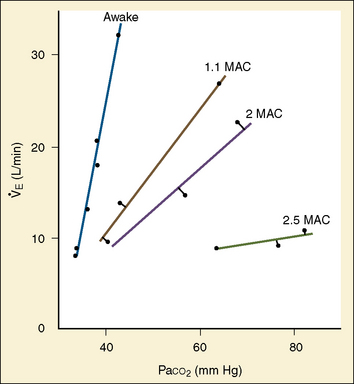
The graphic demonstration of relations between the alveolar or arterial Pco2 and the minute ventilation (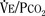 ) is commonly known as the CO2 response curve (Fig. 3-8). This curve normally reflects the response of the chemoreceptors and respiratory center to CO2. The CO2 response curve is a useful means for evaluation of the chemical control of breathing, provided that the mechanical properties of the respiratory system, including the neuromuscular transmission, respiratory muscles, thorax, and lungs, are intact. In normal persons, ventilation increases more or less linearly as the inspired concentration of carbon dioxide increases up to 9% to 10%, above which ventilation starts to decrease (Dripps and Comroe, 1947). Under hypoxemic conditions the CO2 response is potentiated, primarily via carotid body stimulation, resulting in a shift to the left of the CO2 response curve (Fig. 3-8) (Nielsen and Smith, 1951). On the other hand, anesthetics, opioids, and barbiturates in general depress the medullary chemoreceptors and, by decreasing the slope, shift the CO2 response curve progressively to the right as the anesthetic concentration increases (Fig. 3-9) (Munson et al., 1966).
) is commonly known as the CO2 response curve (Fig. 3-8). This curve normally reflects the response of the chemoreceptors and respiratory center to CO2. The CO2 response curve is a useful means for evaluation of the chemical control of breathing, provided that the mechanical properties of the respiratory system, including the neuromuscular transmission, respiratory muscles, thorax, and lungs, are intact. In normal persons, ventilation increases more or less linearly as the inspired concentration of carbon dioxide increases up to 9% to 10%, above which ventilation starts to decrease (Dripps and Comroe, 1947). Under hypoxemic conditions the CO2 response is potentiated, primarily via carotid body stimulation, resulting in a shift to the left of the CO2 response curve (Fig. 3-8) (Nielsen and Smith, 1951). On the other hand, anesthetics, opioids, and barbiturates in general depress the medullary chemoreceptors and, by decreasing the slope, shift the CO2 response curve progressively to the right as the anesthetic concentration increases (Fig. 3-9) (Munson et al., 1966).
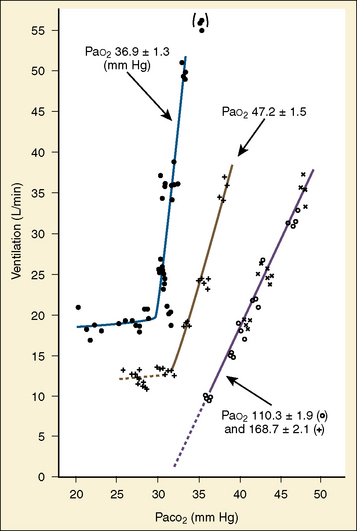
A shift to the right of the CO2 response curve in an awake human may be caused by decreased chemoreceptor sensitivity to CO2, as seen in patients whose carotid bodies had been destroyed (Wade et al., 1970). It may also be caused by lung disease and resultant mechanical failure to increase ventilation despite intact neuronal response to carbon dioxide. In patients with various central nervous system dysfunctions, the CO2 response may be partially or completely lost (Ondine’s curse) (Severinghaus and Mitchell, 1962). In the awake state, these patients have chronic hypoventilation but can breathe on command. During sleep, they further hypoventilate or become apneic to the point of CO2 narcosis and death unless mechanically ventilated or implanted with a phrenic pacemaker (Glenn et al., 1973).
It has been difficult to separate the neuronal component from the mechanical failure of the lungs and thorax, because the two factors often coexist in patients with chronic lung diseases (Guz et al., 1970). Whitelaw and others (1975) demonstrated that occlusion pressure at 0.1 second (P0.1, or the negative mouth pressure generated by inspiratory effort against airway occlusion at FRC) correlates well with neuronal (phrenic) discharges but is uninfluenced by mechanical properties of the lungs and thorax. The occlusion pressure is a useful means for the clinical evaluation of the ventilatory drive.
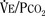 ). Such a change has been interpreted as “a synergistic (or multiplicative) effect” of the stimulus, whereas a parallel shift of the curve has been considered as “an additive effect.” This analysis may be useful for descriptive purposes, but it is misleading. Because ventilation is the product of tidal volume and frequency (
). Such a change has been interpreted as “a synergistic (or multiplicative) effect” of the stimulus, whereas a parallel shift of the curve has been considered as “an additive effect.” This analysis may be useful for descriptive purposes, but it is misleading. Because ventilation is the product of tidal volume and frequency (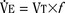 ), an additive effect on its components could result in a change in the slope of the CO2 response curve. Obviously, the responses of tidal volume and frequency to CO2 should be examined separately to understand the effect of various respiratory stimulants and depressants.
), an additive effect on its components could result in a change in the slope of the CO2 response curve. Obviously, the responses of tidal volume and frequency to CO2 should be examined separately to understand the effect of various respiratory stimulants and depressants.Milic-Emili and Grunstein (1975) proposed that ventilatory response to CO2 be analyzed in terms of the mean inspiratory flow (Vt/Ti, where Vt is tidal volume and Ti is the inspiratory time) and in terms of the ratio of inspiratory time to total ventilatory cycle duration or respiratory duty cycle (Ti/Ttot) (Fig. 3-10). Because the tidal volume is equal to Vt/Ti × Ti and respiratory frequency (f) is I/Ttot, ventilation can be expressed as follows:
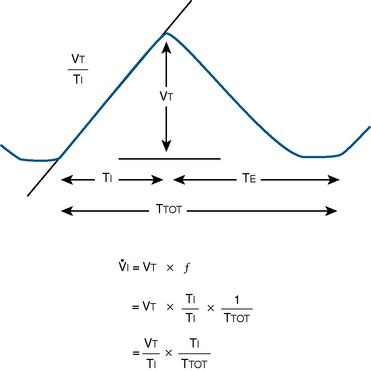
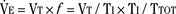
The advantage of analyzing the ventilatory response in this fashion is that Vt/Ti is an index of inspiratory drive, which is independent of the timing element. The tidal volume, on the other hand, is time dependent, because it is (Vt/Ti) × TI. The second parameter, Ti/Ttot, is a dimensionless index of effective respiratory timing (respiratory duty cycle) that is determined by the vagal afferent or central inspiratory off-switch mechanism or by both (Bradley et al., 1975). From this equation, it is apparent that in respiratory disease or under anesthesia, changes in pulmonary ventilation may result from a change in Vt/Ti, Ti/Ttot, or both. A reduction in Ti/Ttot indicates that the relative duration of inspiration decreased or that expiration increased. Such a reduction in the Ti/Ttot ratio may result from changes in central or peripheral mechanisms. In contrast, a reduction in Vt/Ti may indicate a decrease in the medullary inspiratory drive or neuromuscular transmission or an increase in inspiratory impedance (i.e., increased flow resistance, decreased compliance, or both). By relating the mouth occlusion pressure to Vt/Ti, it becomes clinically possible to determine whether changes in the mechanics of the respiratory system contribute to the reduction in Vt/Ti (Milic-Emili, 1977).
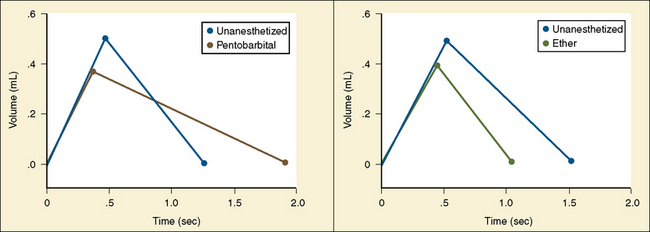
Analysis of inspiratory and expiratory durations provides useful information on the mechanism of anesthetic effects on ventilation. Figure 3-11 illustrates the effect of pentobarbital, which depresses minute ventilation, and diethyl ether, which “stimulates” ventilation in newborn rabbits. With both anesthetics the mean inspiratory flow (Vt/Ti) did not change, but Vt decreased because Ti was shortened. With pentobarbital, however, Te was prolonged disproportionately, and Ti/Ttot and frequency decreased; consequently, minute ventilation was decreased. With ether, on the other hand, ventilation increased as the result of disproportionate decrease in Te and consequent increases in Ti/Ttot and frequency (Milic-Emili, 1977).
Control of Breathing in Neonates and Infants
Response to Hypoxemia in Infants
During the first 2 to 3 weeks of age, both full-term and premature infants in a warm environment respond to hypoxemia (15% oxygen) with a transient increase in ventilation followed by sustained ventilatory depression (Brady and Ceruti, 1966; Rigatto and Brady, 1972a, 1972b; Rigatto et al., 1975a) (Fig. 3-12). In infants born at 32 to 37 weeks’ gestation, the initial period of transient hyperpnea is abolished in a cool environment, indicating the importance of maintaining a neutral thermal environment (Cross and Oppe, 1952; Ceruti, 1966; Perlstein et al., 1970







Full access? Get Clinical Tree


