Dimensions of the pain experience
Pain has a protective function and may or may not be associated with tissue damage.
Pain should not be equated to nociception. Nociception is often a component of pain symptoms but not necessarily so.
Intrinsic modulatory mechanisms exist in the body which attenuate the intensity of the pain experience.
Sensitisation mechanisms exist which intensify pain symptoms, resulting in the phenomenon of hyperalgesia.
Pain possesses a subjective and affective element as a result of connections between the pain system, cortical centres and the limbic system.
Pain levels are also influenced by past experience and anticipation due to interaction between the pain system and the prefrontal cortex.
Pain can affect visceral and neuroendocrine function as a result of interconnections with medullary centres and the hypothalamus.
Nociceptors
The primary afferent neurones for pain are referred to as ‘nociceptors’, which possess specialised nerve endings in almost all tissues of the body. These primary afferents respond to different types of noxious stimuli. Such stimuli are most commonly mechanical, thermal or chemical.
Nociceptor characteristics
The type of stimuli they are sensitive to
The class of nerve fibre serving the receptor
The receptor response characteristics – slow or fast, short or long latency
The ion channel proteins expressed on the receptor cell membrane
Unimodal receptors respond to mechanical distortion, thermal receptors to heat, and polymodal receptors to mechanical distortion, heat, cold or chemical stimuli. The majority of receptors are polymodal, being sensitive to mechano-heat (MH) stimuli. Some receptors are only sensitive to mechanical stimuli and are referred to as mechanically sensitive afferents (MSA) while others are insensitive to mechanical stimuli (mechanically insensitive afferents, MIA).
There are two main types of nociceptor fibre which transmit nociceptor signals to the central nervous system, C fibres and Aδ fibres.
C nociceptors are unmyelinated, with conduction velocities < 2 m s−1. They evoke slow-response burning pain and dull aching pain. They are mostly polymodal, responding to heat, mechanical and chemical stimuli. These nociceptors may be referred to using the nomenclature CMH (C mechano-heat nociceptors). Some C-fibre nociceptors are mechanically insensitive afferents, and are referred to as C-fibre MIAs.
Aδ nociceptors are connected centrally by myelinated fibres with conduction velocities between 15 and 55 m s−1. Aδ fibres evoke sharp fast-response pains of a burning, stabbing, pricking and aching nature. There are two types of Aδ nociceptor. Type I have higher conduction velocities (> 25 m s−1) and are found in hairy and glabrous skin (palms of hands and soles of feet). These Aδ nociceptors may be referred to as AMHs (A mechano-heat nociceptors). Type II are slower, with conduction velocities of around 15 m s−1, and are distributed in hairy skin only. An outline of their contrasting responses to stimuli is shown in Figure 19.2.
| Response | Type I | Type II |
|---|---|---|
| Mechanical | Usually sensitive (MSA) | Usually insensitive (MIA) |
| Heat | Insensitive (high threshold) initially with increasing response to prolonged stimulus (> 5 s) | Sensitive (low threshold) initially with decreasing response to prolonged stimulus |
| Long latency | Short latency | |
| Chemical | Sensitive | Sensitive |
Cellular functions of nociceptors
Nociceptor functions
Transducing and encoding noxious stimuli to generate afferent action potentials
Synthesising and releasing neuropeptides such as calcitonin gene-related peptide (CGRP) and substance P, both peripherally and centrally
Synthesising and releasing neurotrophins such as glial cell-line-derived neurotrophic factor (GDNF) and nerve growth factor (NGF), which modulate gene expression in the nociceptor
Synthesising and releasing neurotransmitters at the primary synapse, e.g. glutamate
Synthesising and releasing inflammatory mediators which sensitise the nociceptive pathway centrally and peripherally
Nociceptor ion channels
Nociceptors possess families of ion channels in their cell membranes which initiate their functions and have the following properties:
They are usually specific for sodium, potassium, calcium or hydrogen ions (acid-sensing ion channels, ASICs). However, non-specific ion channels also exist.
Transient receptor potential (TRP) channels in the nociceptor membrane transduce noxious physical or chemical stimuli to initiate action potentials.
TRP channels can be sensitised or activated by inflammatory mediators released from damaged tissue or by the nociceptors themselves. This results in hyperalgesia.
Voltage-gated channels are opened at different thresholds of membrane potential. Such channels can be triggered by the transient changes in membrane potential produced when TRP channels are activated. TRP channels can thus initiate an action potential. In this way a stimulus can be encoded into action potentials with a rate which varies according to the intensity of the stimulus.
Up to nine different voltage-gated sodium channels have been identified in a single nociceptor cell membrane. Individual members of such families of ion channels are identified by established nomenclature: e.g. ion channels specific to nociceptors have been identifed as Nav 1.8 and 1.9 (Na, sodium; v, voltage-gated; ‘1’ gene subfamily; ‘8’ isoform).
Nociceptor functions are initiated by ion channels and receptors in the nociceptor cell membrane. Each nociceptor possesses different types of ion channel. They include transient receptor potential (TRP) ion channels, ligand-gated ion channels and voltage-gated ion channels.
Nociception and inflammatory mediators
When a noxious stimulus is applied, chemical mediators such as prostaglandins, hydrogen ions and kinins are released by the nociceptor or as a result of tissue damage. These substances not only initiate nociception, but also produce hyperalgesia. In addition, these agents mediate the inflammatory process by inducing the following changes:
Increases in local blood flow and vascular permeability
Activation and migration of immune cells
Release of growth and trophic factors from surrounding tissues
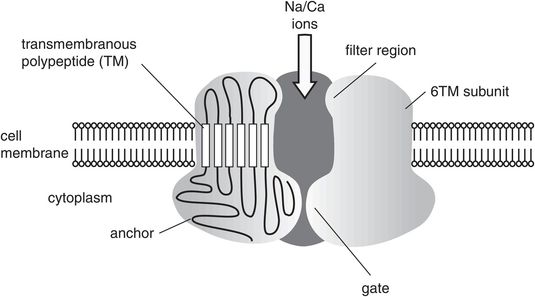
Transient receptor potential (TRP) channels
TRP channels are the ‘transducer’ channels in the plasma membranes of a wide variety of animal and human cells (Figure 19.3).
They are activated by different physical and chemical stimuli (pressure, temperature, osmotic pressure, vibration, H+, capsaicin, cannabinoids).
When activated, the TRP channels produce a transient drop in resting membrane potential to trigger action potentials and thus mediate sensory modes such as heat and pressure sensation, vibration sense and pain.
At least seven subfamilies of TRP channel have been identified.
A prominent TRP channel in hyperalgesic conditions is the TRPV1 (vanilloid 1) channel, which is a calcium/sodium channel. The TRPV1 channel can also be potentiated by heat and H+, and is inhibited by glycine or intracellular phosphatidylinositol biphosphate (PIP2).
Transient receptor potential (TRP) channel
There are different mechanisms by which inflammatory mediators influence nociceptor function:
Activation of membrane ion channels, e.g. lipid metabolite activation of TRPV1 channels, proton activation of ASICs, ATP activation of purinergic (P2X) channels.
Activation of G-protein-coupled receptors which increase the activities of second messengers such as cyclic adenosine monophosphate (cAMP) or inositol triphosphate (IP3). These in turn may lead to structural changes in membrane ion channels or changes in the activity of intracellular enzymes affecting membrane excitability.
Activation of cytokine receptors by interleukins (IL-1) or tumour necrosis factor α (TNF-α).
Activation of tyrosine kinase receptors TrkA, TrkB, by specific neurotrophic factors such as NGF (which activates TrkA), and brain-derived neurotrophic factor (BDNF) (which activates TrkB)
Some examples of inflammatory mediators and their origins are shown in Figure 19.4.
| Mediator | Source/origin |
|---|---|
| Bradykinin | Damaged tissue |
| Arachidonites | Arachidonic acid |
| Serotonin | Platelet and mast cells |
| Histamine | Mast cells |
| Cytokines (TNF-α, IL-1) | Immune system |
| Intracellular molecules (ATP, NO) | Damaged tissue |
| Neurotrophins (NGF, GDNF, BDNF) | Nociceptors |
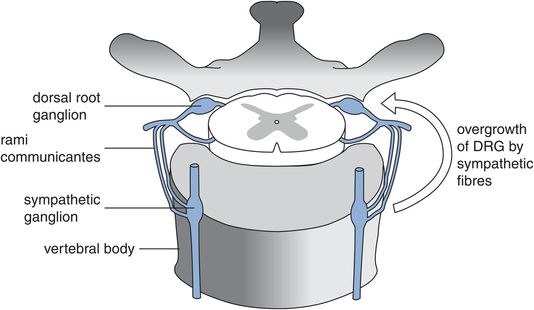
The dorsal root ganglion
The dorsal root ganglion (DRG) contains the cell bodies for all primary afferents including the nociceptors. The primary afferent fibres are classified according to conduction velocity. Aα and Aβ fibres are the fastest group with large cell bodies, forming about 40% of the DRG cells. Aδ and C fibres are slower with small cell bodies, and account for about 60% of the DRG population. Nociceptors form the majority of the small cells. They can be further classified into peptidergic if they contain peptides (CGRP, substance P, somatostatin) or non-peptidergic. Approximately 50% of C and 20% of Aδ fibres are peptidergic. Although nociceptors are mainly small-cell-bodied, some nociceptors also exist in the faster larger-neurone groups. Specific populations of DRG neurones can be identified by the expression of different neurotrophin receptors on their cell membranes.
The DRG is situated in the lateral foramen between neighbouring vertebral bodies and is in close relationship to the sympathetic chain (Figure 19.5). This anatomical relationship becomes significant in chronic pain conditions such as complex regional pain syndrome (CRPS), in which abnormal overgrowth of the DRG by fibres from the sympathetic chain is thought to be responsible for the sympathetically maintained symptoms and trophic changes in the affected areas.
The dorsal horn
Nociceptors terminate almost exclusively in the dorsal horn of the spinal cord. The grey matter of the spinal cord was divided into 10 laminae (numbered I to X) by Rexed in 1952. The dorsal horn is composed of laminae I to VI (Figure 19.6). The majority of nociceptor fibres (Aδ and C) terminate in two areas, which are laminae I/II (marginal layer and substantia gelatinosa) and lamina V. It should be noted that in addition to Aδ and C fibres, a small fraction of Aβ afferent fibres are nociceptive with high-threshold mechano-heat characteristics. The nociceptive fibres synapse with second-order neurones or interneurones within the dorsal horn.
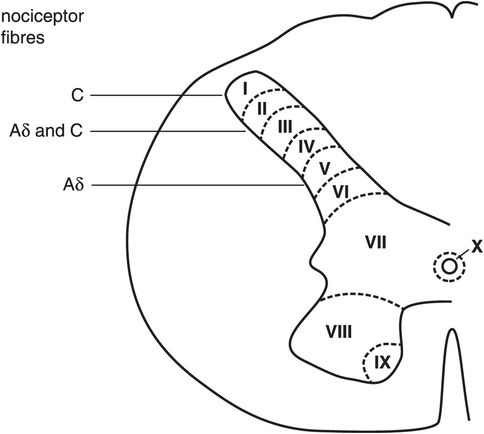
Dorsal horn laminae in spinal cord
The second-order neurones in the dorsal horn may be excited or inhibited by different configurations of the nociceptors and interneurones. Sensitisation of the primary synapse by the descending modulatory system or interneurones can also occur in hyperalgesic states (Figure 19.7). Similarly, various configurations of the descending modulatory system and interneurones may occur to inhibit transmission in the primary synapse (Figure 19.8).
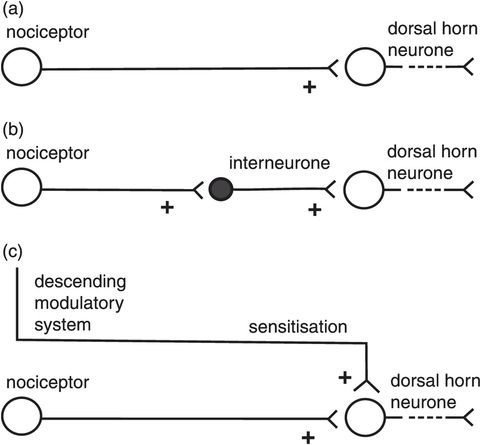
Excitatory connections in the dorsal horn
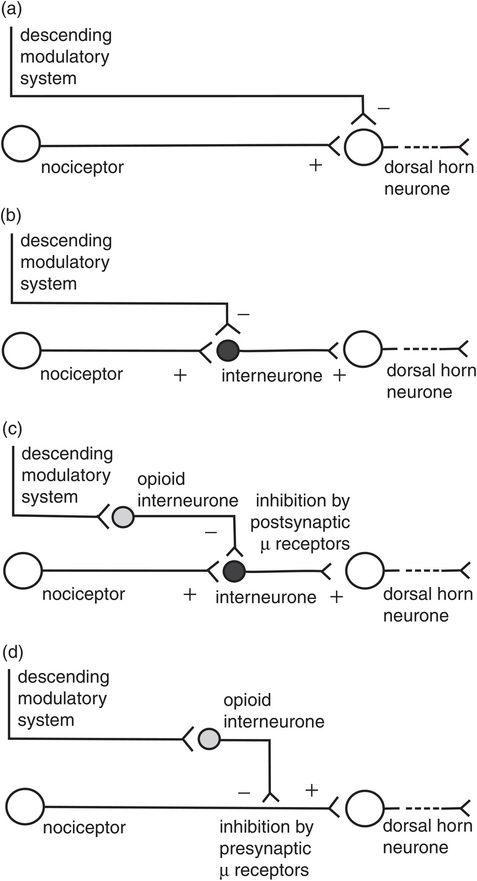
Inhibitory connections in the dorsal horn
The dorsal horn contains the following neuronal elements which participate in nociception:
Primary afferent axons and their synapses connecting with secondary neurones or interneurones
Secondary neurones which project afferent information to the brain via ascending tracts
Interneurones which form interconnection circuits at one level, or can ascend and descend to interconnect in other segments
Descending axons from supraspinal centres modulating nociception
Interneurones
The great majority of neurones in the dorsal horn are interneurones. Inhibitory and excitatory interneurones exist. Excitatory interneurones release glutamate as the neurotransmitter into the primary synapse. Inhibitory neurones use γ-aminobutyric acid (GABA) and glycine as their neurotransmitters, and may be stimulated by primary afferents, the descending modulatory system or non-nociceptive primary afferents such as Aβ fibres. The interneurones form interconnection circuits which participate in the following aspects of dorsal horn activity:
Gating of nociceptive signals (see The ‘gate control’ theory, below)
Sensitisation of the primary synapse
Presynaptic inhibition of primary afferents
Relaying primary afferent signals to the appropriate ascending tracts
The primary synapse
Nociceptors are the primary afferent neurones of the pain system; they stimulate action potentials in the dorsal horn neurone via the primary synapse. The primary synapse is central to the mechanism by which the pain system controls its sensitivity. The main excitatory neurotransmitter in the primary synapse is glutamate (Figure 19.9).
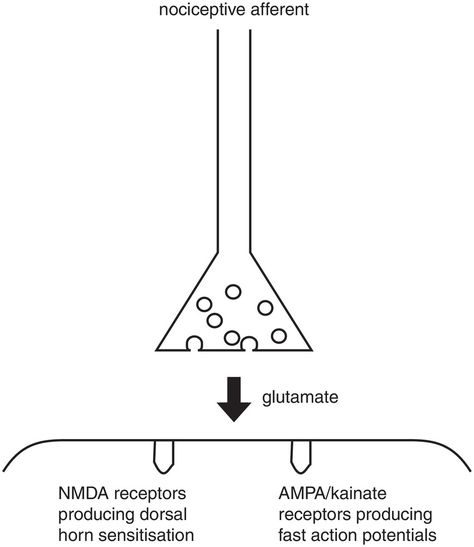
Excitation at the primary synapse
Primary afferents can also inhibit action potentials in dorsal horn neurones by stimulating inhibitory interneurones. The main inhibitory neurotransmitters are GABA and glycine (Figure 19.10).
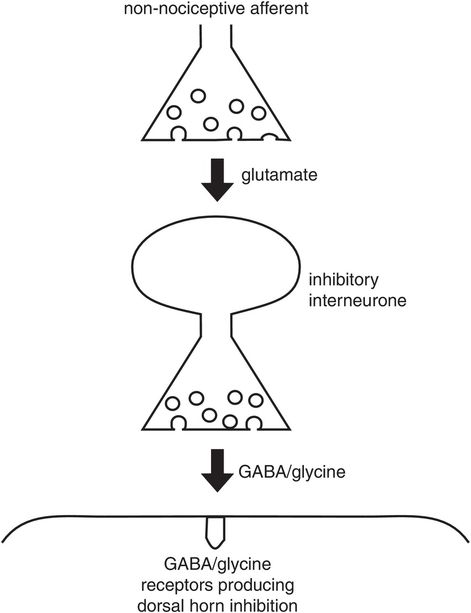
Postsynaptic inhibition at the primary synapse
Synaptic transmission is also influenced by a variety of neuromodulators which act through a spectrum of pre- and postsynaptic membrane receptors. These modulators include endogenous opioids, which suppress transmission, but other modulators can enhance transmission. Neuromodulators in the pain system are described further below.
Descending control of the primary synapse is exerted by supraspinal centres which form the descending modulatory system. This system not only can inhibit synaptic transmission but may also facilitate it, leading to sensitisation.
Control of synaptic transmission in the dorsal horn is therefore a result of complex interactions between afferent, descending, ascending and interneurone fibres (Figures 19.7 and 19.8).
Neurotransmitters in the pain system
Many different synapses exist in the pain pathways of the CNS. It is recognised that each synapse operates with multiple transmitters. Some examples of known neurotransmitters in the pain system are presented in Figure 19.11.
| Neurotransmitter | Location | Comments |
|---|---|---|
| Excitatory neurotransmitters | ||
| Glutamate | Widespread throughout CNS. Primary afferents | Main excitatory transmitter in CNS. > 50% of DRG neurones. The main excitatory transmitter in nociception, acting via ionotropic receptors (AMPA, NMDA and kainite receptors) |
| Substance P | Primary afferents | 30% of DRG neurones, mainly small sensory afferents. Small peptide agonist for neurokinin (NK1) receptor. Excitatory role in ‘wind up’ |
| Calcitonin gene-related peptide (CGRP) | Dorsal root ganglion cells | 50% of DRG neurones, both small and large pain fibres. Released by thermal and mechanical stimuli. Excitatory role in ‘wind up’ |
| Inhibitory neurotransmitters | ||
| γ-Aminobutyric acid (GABA)/glycine | Descending modulation system, dorsal and ventral horn | Potent inhibition of spinal cord neurones. Act through ligand-gated channels |
| Noradrenaline | Dorsal horn and descending modulation system | Dorsal horn inhibition |
| Serotonin | Dorsal horn and descending modulation system | Dorsal horn inhibition |
| Glutamate | Dorsal horn | Inhibitory effects in dorsal horn exerted via metabotropic receptors |
Receptors in the primary synapse
The presynaptic and postsynaptic membranes of the primary synapse possess different families of receptor proteins (Figure 19.12). Some receptors (ionotropic) control associated ion channels. Others, such as tyrosine kinase (Trk) receptors, signal intracellular pathways, resulting in the synthesis and release of peptides, neurotransmitters or neurotrophic factors. These are metabotropic receptors.
| Receptor | Location | Comments |
|---|---|---|
| NMDA | Postsynaptic membrane | Important in producing long-lasting sensitisation by allowing calcium influx into the projecting dorsal horn neurones. Normally suppressed by Mg 2+, may also be blocked by ketamine |
| AMPA | Postsynaptic membrane | Produce fast excitatory postsynaptic action potentials. The main receptor responsible for transmission between primary afferents and dorsal horn neurones. Mainly sodium ionophores |
| Kainate | Postsynaptic membrane | Produce slow postsynaptic potentials associated with activation of high-threshold primary afferents. Sodium and potassium ionophores |
| NK1 | Metabotropic receptors activated by substance P. Activate nociceptive dorsal horn neurones and produce hyperalgesia. Increase intracellular Ca2+ | |
| CGRP receptors | Postsynaptic membrane | Produce facilitation of the dorsal horn response |
| Opioid δ, κ, μ | Postsynaptic and presynaptic membranes | Inhibition of dorsal horn neurones |
| GABAA, GABAB | Postsynaptic membrane | Inhibition of dorsal horn neurones GABAA increases Cl− conductance and stabilises postsynaptic membrane potential GABAB decreases Ca2+ conductance and causes postsynaptic membrane hyperpolarisation |
| α2 | Presynaptic membrane | Selective inhibition of nociceptive primary afferents leading to analgesia |
| Serotonin receptors | Postsynaptic membrane, NMDA receptors | Inhibition of dorsal horn transmission. Some serotonin receptors associated with excitatory action |
Primary synapse receptors may be classified into metabotropic receptors and ionotropic receptors:
Metabotropic receptors activate intracellular pathways when triggered by an agonist.
Ionotropic receptors alter the conductivity of an associated ion channel.
The main excitatory transmitter is glutamate, which activates three different subtypes of ionotropic receptor on the postsynaptic membrane. These are NMDA (N-methyl-D-aspartate), AMPA (α-amino 3-hydroxy 5-methyl 4-isoxazolepropionic acid) and kainate receptors (Figure 19.9). The NMDA receptor has a prominent role in chronic pain conditions, while the AMPA receptor mediates the transmission of fast action potentials.
In addition to these ionotropic receptors there are modulatory receptors on the postsynaptic membrane which can facilitate or inhibit action potential formation.
The main inhibitory receptors are γ-aminobutyric acid (GABA) and glycine receptors (Figure 19.8). Other inhibitory receptors include μ and κ opioid receptors and adenosine receptors, which are G-protein coupled.
The presynaptic nociceptor membrane has a range of receptors which include opioid (μ, δ and κ), nicotinic and muscarinic receptors. These also function to modulate synaptic transmission (Figure 19.13).
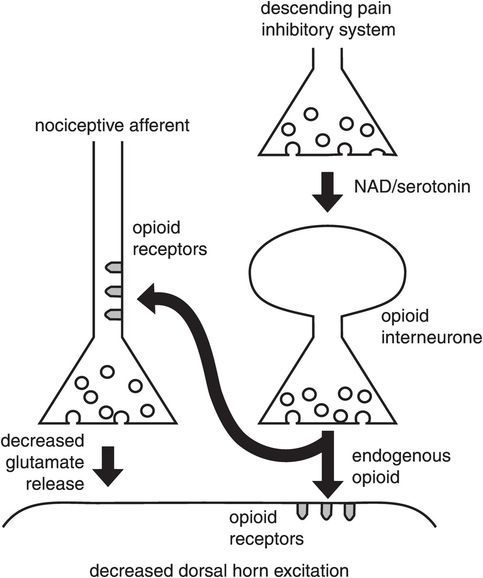
Presynaptic inhibition at the primary synapse
Related posts:






Full access? Get Clinical Tree


